Гены,
находящиеся в разных локусах также могут влиять друг на друга. При этом можно
выделить несколько типов подобного взаимодействия. Гены,
находящиеся в разных локусах также могут влиять друг на друга. При этом можно
выделить несколько типов подобного взаимодействия.
Гены,
не проявляющие собственного действия, но усиливающие или ослабляющие эффект
действия других генов, называются генами-модификаторами. Изучение
окраски у млекопитающих показало, что наряду с крайними формами, обладающими
полным развитием пигмента или его отсутствием, наблюдается целый ряд генотипически
обусловленных форм. Так, белая пятнистость у собак варьирует от нескольких
белых волосков на месте первичной точки депигментации до полностью белой собаки
с небольшим пучком окрашенных волос в одном из пигментных центров. В пределах
генотипа, определяемого локусом белой пятнистости S, можно выделить
массу переходных форм.

Разные
варианты белой пятнистости собак
Широкую
вариабельность, обусловленную генами-модификаторами, имеют чепрачный и тигровый
окрасы собак.
Если
признак образуется при наличии двух пар неаллельных генов, дающих при
совместном действии не тот эффект, который каждый из них обусловливает
самостоятельно, то такие аллели обозначают как комплементарные
(дополняющие друг друга). В качестве примера комплементарности у собак обычно
приводят взаимодействие генов локусов В и Е, определяющих
окраску.
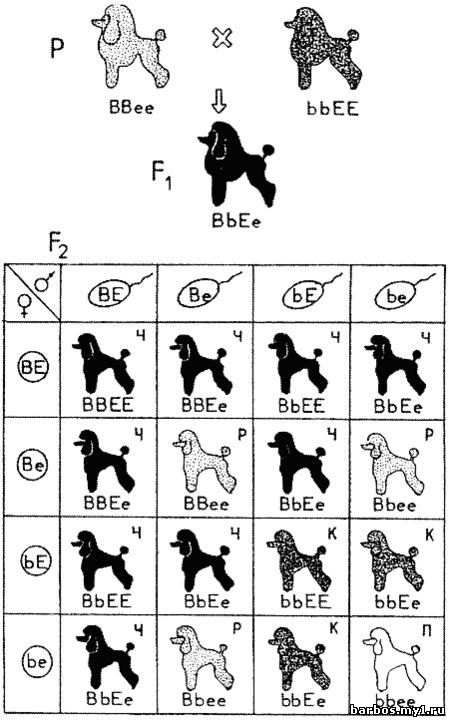
Комплементарное
взаимодействие генов, определяющих окраску собак: соотношение — 9 черных (Ч): 3
коричневых (К): 3 рыжих (Р): 1 палевая (П)
Гены
локуса В отвечают за синтез черного ( В) или коричневого ( b)
пигмента. Гены локуса Е отвечают за распределение этих пигментов. Аллель
Е способствует распространению черного или коричневого пигмента по всему
телу собаки. Аллель е препятствуют их синтезу в шерстном покрове. Собаки
с генотипом ее — рыжие или желтые. Черный или коричневый пигмент при
этом сосредоточен только на кожных покровах морды собаки.
Формирование
окраски собаки зависит от присутствия обоих пар генов. Собаки с генотипом ЕЕ
или Ее — черные или коричневые в зависимости от аллелей B или b.
При Е-ВВ или Е-Bb — собака черная, при E-bb — коричневая.
Собаки
с генотипом ееВ- — рыжие с черным носом. Собаки генотипа bbee —
обычно палевые или светло-желтые со светлым носом.
Комплементарным
взаимодействием генов обусловлен особый тип паралича задних конечностей у
помесей датского дога с сенбернаром. Генетический анализ проведенный Стокардом
(Stockard, 1936), показал, что при чистопородном разведении как датских догов,
так и сенбернаров паралич не развивается.
Аналогичное
заболевание отмечалось у некоторых помесных собак-ищеек (Петухов и др., 1985).
В
любой паре аллелей доминантный ген препятствует (полностью или частично)
проявлению своего рецессивного партнера. Но иногда действие доминантного аллеля
подавляется действием гена из другого локуса. Всемогущий ген, который блокирует
действие другого гена или генов называется эпистатичным. А само явление
— эпистазом. Гены, действие которых подавлено, называется
гипостатичными.
Так,
рецессивные гены окраски собак из локуса C не позволяют синтезироваться
пигментам, определяющим окраску шерсти. Собака гомозиготная по ним — белая.
Один
и тот же мутантный признак может проявляться у одних и не проявляться у других
особей родственной группы. Способность данного гена проявлять себя
фенотипически называется пенетрантностью. Пенетрантность определяется по
проценту особей в популяции, имеющих мутантный фенотип. При полной
пенетрантности (100%) мунантный ген проявляет свое действие у каждой особи. При
неполной пенетрантности (меньше 100%) ген проявляется фенотипически не у всех
особей.
У
собак достаточно часто встречаются видоизменения хвостов в виде их
укороченности, разнообразных изломов и изгибов. Можно предположить, что
многообразие этого признака обусловлено его неполной пенетрантностью.
Степень
пенетрантности может сильно изменяться под воздействием условий среды.
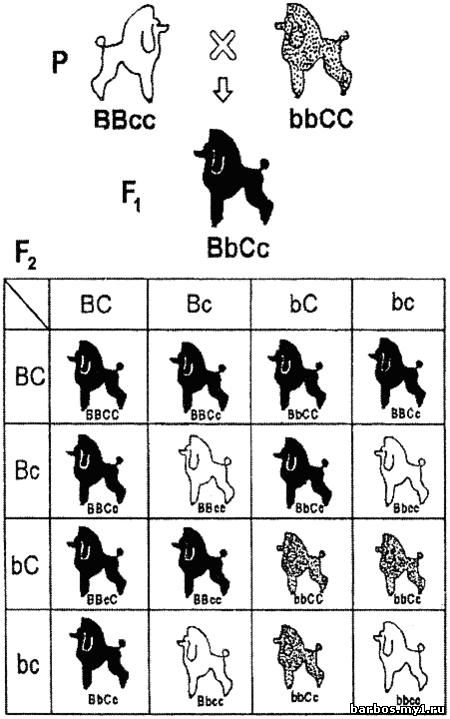
Схема
дигибридного расщепления с рецессивным эпистазом: вВ F 2 получено 9 черных: 3
коричневых: 4 белых собаки. Таким образом, наблюдается характерное для
рецессивного эпистаза отклонение от теоретически ожидаемого расщепления 9:3:3:1
Часто
особи, обладающие тем же генотипом в отношении какого-либо наследственного
признака, очень сильно различаются по его экспрессивности, то есть
степени проявления данного признака. Один и тот же ген у разных особей в
зависимости от влияния генов-модификаторов и внешней среды может проявить себя
фенотипически по-разному. Внешняя среда и гены-модификаторы могут изменить экспрессию
гена, то есть выражение признака.
В
отличие от пенетрантности, которая указывает, у какой доли особей в популяции
проявляется данный признак, экспрессивность относится к изменчивости признака у
тех особей, у которых он проявляется. Так у собак экспрессивность развития
прибылых пальцев варьирует от полностью развитых пальцев на обеих задних
конечностях до наличия их в зачаточном состоянии только на одной конечности.
Подобная вариация экспрессивности характерна и для других наследуемых
признаков, в частности и для вышеупомянутых хвостов.
Экспрессивность
и пенетрантность гена в значительной степени зависят, по-видимому, от влияния
генов-модификаторов и условий развития особей.
Достаточно
широко распространено явление плейотропии — влияния одного гена на
развитие двух и более признаков. Классический «собачий» пример плейотропного
влияния — действие фактора Мерля, (локус М; окраса собак). Аллель
M в гетерозиготном состоянии Мm дает пятнистость типа «арлекин»
характерную для догов. Аллель M в гетерозиготе Mm в сочетании с
подпалом дает «мраморный» (blue-merle) окрас, типичный для колли и шелти. В
гомозиготном состоянии ММ он ведет к рождению чисто белых щенков (white-merle)
со значительными аномалиями органов чувств. Такие щенки часто погибают еще до
рождения, а если и рождаются живыми, то их жизнеспособность резко снижена.
Явление
плейотропии объясняется тем, что гены плейотропного действия контролируют
синтез ферментов, участвующих в многочисленных обменных процессах в клетке и в
организме в целом и, тем самым, одновременно влияющих на проявление и развитие
многих признаков.
Некоторые
гены вызывают столь сильные отклонения от нормы, что снижают жизнеспособность
организма или даже приводят его к гибели. Такие гены называют летальными,
то есть смертоносными, или сублетальными — снижающими жизнеспособность.
В большинстве случаев летальные гены полностью рецессивны, поэтому
гетерозиготные носители этих генов фенотипически совершенно неотличимы от
нормальных особей. В гомозиготном состоянии такие гены могут нарушать
нормальный ход развития эмбриона на любых стадиях. О возможности наличия
летальных генов можно косвенно судить по снижению средней численности пометов
или по выпадению некоторой ожидаемой части фенотипов в расщеплении.
Так
в случае скрещивания упоминавшихся выше черно-мраморных собак, гетерозиготных
по фактору Мерля получается вместо ожидаемого соотношения 3:1,
получается 2:1 т. е. 2 мраморных и 1 черная собаки Mm × Mm
= ММ: 2Mm: mm, где ММ белая нежизнеспособная собака. Белые
щенки чаще не рождаются вовсе, так как погибают задолго до рождения.
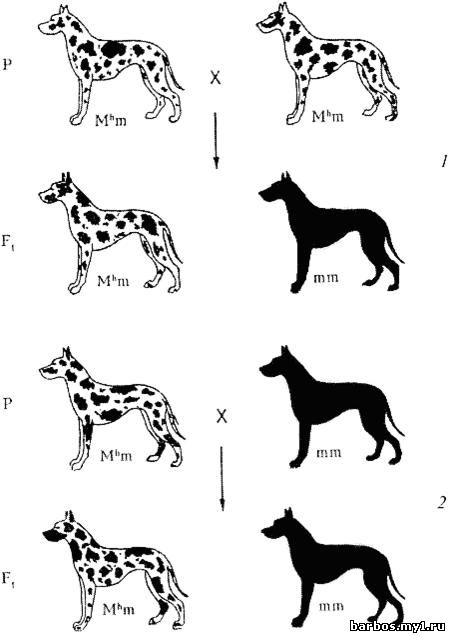
Наследование
мраморности типа «арлекин» у догов. Ген М h (фактор Мерля) — доминантный с
рецессивным летальным действием: 1 — скрещивание мраморных догов между собой; 2
— анализирующее скрещивание
Фактор
Мерля относится к категории доминантных летальных генов, которых значительно
меньше, чем рецессивных. При желании его носителей легко изъять из разведения,
так как они имеют характерный фенотип. Одни летальные гены обуславливают
крупные аномалии, другие — нарушения физиологических процессов. Пути
губительного воздействия большинства летальных генов не ясны. Таких генов может
существовать сколь угодно много. Показано, что каждый человек является
носителем в среднем 4–9 «вредных» или летальных генов. Аналогичных результатов
можно ждать и по собакам. Известны летальные гены, которые при проявлении в
эмбриональном состоянии, опасны и для жизни беременной суки, например, при
наследственной контрактуре мышц плода, вследствие чего сука не может
разродиться.
Взаимодействие
генов, когда при их сочетании в одном организме развивается совершенно новая
форма признака называется новообразованием.
Иногда
новообразования приводят к появлению признаков дикого фенотипа. В этом случае
их называют атавизмами, то есть возвратом к предковой форме или
возвратом в дикий тип.
Частичные
возвраты в дикий тип возможны при скрещивании двух особей одной породы, если
эти производители происходят из удаленных друг от друга неродственных
популяций. По-видимому, подобным образом можно объяснить и большое сходство
между дворнягами, обитающими в самых разных местах.
Признаки,
сцепленные с полом
Признаками, сцепленными с полом
называют такие, которые формируются под воздействием генов, локализованных в Х-хромосоме.
Наиболее характерным примером наследования признака сцепленного с полом
является наследование гемофилии собак. У собак больных гемофилией, в крови не
хватает фактора, который, взаимодействуя с кровяными пластинками
(тромбоцитами), ускоряет превращение протромбина в тромбин. Гемофилия собак
аналогична гемофилии человека и также обусловлена сцепленным с полом
рецессивным геном. Ген, определяющий развитие гемофилии находится в Х-хромосоме
и является рецессивным по отношению в нормальному аллелю. Следовательно,
гемофилия проявляется только у гомозиготных самок (несущих этот ген в обеих Х-хромосомах)
и гемизиготных самцов, несущих ген гемофилии в Х-хромосоме.
Щенки-гемофилики обычно погибают в раннем возрасте от наружных или внутренних
кровотечений. Сохранить до половозрелого состояния такого самца возможно только
при постоянном введении специфических препаратов. Самки же неминуемо гибнут не
позже первой течки. Гетерозиготные самки внешне абсолютно нормальны и
плодовиты. Однако половина их детенышей мужского пола страдает гемофилией и
половина детенышей-самок оказывается гетерозиготными по этому гену.
Родители:
сука-носительница
гена гемофилии
ХHХh
кобель
нормальный
XHY
—
половая хромосома, несущая нормальный аллель
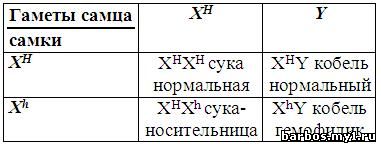
XH
Xh — половая
хромосома, несущая ген гемофилии
К
признакам сцепленным с полом относятся также врожденный гипотрихоз,
отмеченный у такс и карликовых пуделей; мышечная дистрофия у ретриверов;
синдром пошатывания, связанный с гипомиелинизацией и обнаруженный у
чау-чау и ряда других пород; подвывих запястья; а также диафрагмальная
грыжа, описанная у золотистых ретриверов.
Признаки
ограниченные полом
Некоторые
признаки совершенно независимо от места локализации генов их вызывающих,
проявляются только у особей одного пола. Это так называемые ограниченные
полом признаки. Таковы, например, дефекты развития половой системы,
молочность и т. д. Одно из таких явлений — крипторхизм —
непрохождение одного или обоих семенников через паховый канал в мошонку.
Крипторхизм бывает двусторонний, право- или левосторонний и может быть вызван
разными причинами: узостью пахового канала, короткими связками семенников,
недоразвитием семенников. Неопустившиеся семенники могут находиться в разных
местах брюшной полости. Крипторхизм может быть как врожденным, так и
приобретенным. Среди разных его форм существует и генетически обусловленная.
Однако из-за широкой вариабельности этого признака делать однозначные выводы о
его природе невозможно. И уж совершенно неверно трактовать его как моногенный признак,
локализованный в Х-хромосоме.
Закон
гомологических рядов Н.И. Вавилова
Закон
был сформулирован Н.И. Вавиловым в 1920 году. Н.И. Вавилов обнаружил, что все
виды и роды, генетически близкие между собой характеризуются тождественными
рядами наследственной изменчивости.
В
основе закона гомологических рядов лежит параллелизм генотипической
изменчивости у особей со сходным набором генов.
Этот
закон имеет универсальный характер. Обнаружено сходство мутаций у раных видов
животных. Так были отмечены проявления аналогичных форм аномалий у собак,
кошек, кроликов, свиней, человека и др., что указывает на сходство строения
многих ферментов и белков и соответственно на сходство генотипов. Таким
образом, зная формы наследственных изменений у одного вида животных, можно
предполагать, что они имеются или могут возникнуть и у другого близкого вида.
Особенно тщательно изучены наследственные аномалии сельскохозяйственных
животных и человека. У собак описано значительно меньше аномалий, однако это
говорит лишь о меньшей изученности этого вида. Таким образом, при обнаружении
новой аномалии у собак следует поинтересоваться, не описана ли она для других
видов животных.
В
основных породообразующих группах собак наблюдается гомология по многим
признакам. Так, например, длина ног — ахондроплазия встречается у овчарок
(вельшкорги), терьеров (скай терьеров, селихам терьеров, денди даймонт
терьеров), гончих (бассет) легавых, догообразных (бульдоги), собак Тибета
(лхасское апсо, ши тсу), периодически встречаются элементы ахондроплазии у
пуделей. Подобных собак не отмечено только в группе борзых, поскольку этот
признак является противоположным по отношению к акромегалии.
Во
всех породных группах встречаются как гигантские, так и карликовые формы. У
овчарок (комондор — шипперке), догообразных (мастиф — французский бульдог),
терьеров (эрдельтерьер — той терьер), гончих (бладхаунд — бигль), шпицеобразных
(аляска маламут — померанский шпиц), борзых (ирландский волкодав — левретка).
Гомологическая изменчивость в размерах от гигантов до карликов наблюдается и
среди узких породных групп. Например, шнауцеров (ризен — миттель — цверг), такс
(стандартная — карликовая — кроличья), пуделей (стандартный — малый —
карликовый — той пудель).
Признаками,
проявляющими гомологию, являются окрас и тип шерсти. |